原生動物是如何演化成后生動物的?

 溫柔著品察宿世界。
溫柔著品察宿世界。
在本回覆中,你將看到:
Nothing in biology makes sense except in the light of evolution. ——Dobzhansky
在那黑色的生物之海上,進化乃閃爍的常識燈塔。(誤)——杜布贊斯基
還將看到:
答本家兒無邪地借著進化之光往生命史里一瞥,卻像是看到了深不見底的幽暗天坑,旋即心生驚駭,擔憂本身薄弱的根柢撐不起如斯宏偉絢麗的大廈,擔憂本身任何掉察的論述都將當作為對這神圣光景的玷辱。但此次的故事其實過分出色,即便難以做到完整呈現,我也不由得想與列位分享管窺一豹的欣喜與打動。
現實環境是,在閱讀 Nature 近期頒發的文章 從單細胞生物到多細胞生物的進化機制 時,適值看見了這個被提出好久的問題還沒獲得回覆,我就深思著用這篇研究來填上這個浮泛該當綽綽有余,但讀著讀著才發現,本身妄想著只花一點點時候就將這個弘大的、尚未取得共識的議題回覆詳盡,其實是太自不量力了。是的,學界還不確信這個解答是完整的,但不妨,我想更為遍及的歡愉,是每小我都可以或許借著研究者們得出的線索,在這些尚未獲得解決的問題上勾勒出讓本身安心的謎底。
* 有關“演化”與“進化”:都是對英文“evolution”的翻譯,在當下的中文語境中,兩個詞的涵義完全不異。人們對于這兩種翻譯的爭論不在本文的會商規模,為論述便利,本文一律用“進化”。
太長不看版的結論
從現有的證據來看,從單細胞祖先 (long long ago 的原活潑物) 進化出多細胞動物 (后活潑物) 不是一蹴而就的,是漸進且漫長的。這一過程中最主要的是多細胞動物兩個焦點特征的呈現:細胞堆積 (Multicellularity) 與細胞分化 (Differentiation)。
- 多細胞動物細胞群體的形當作是克隆型的,單細胞祖先上發生的有關胞外基質排泄以及胞漿移動調控的突變,在增殖割裂時使本該分手的子代細胞與親代黏連。呈現在多細胞真核生物門類中的另一種堆積體例沒有被動物祖先采用:堆積型——異質的個別在群居糊口中逐漸同化。
- 細胞的邃密分工合作是多細胞生物的特點。在諸多門類的單細胞生物的分歧糊口史期間中,已經呈現了細胞類型、心理功能上的差別,也就是說,細胞成長出形形色色分歧的形態與功能的時候要早于多細胞動物呈現的時候。所以多細胞動物若何進化出細胞分化,關頭不在于若何讓細胞獲得各類“技術”,而在于若何統籌協調利用分歧“技術”的細胞構成完整的個別。現有證據傾標的目的于撐持這樣的結論:多細胞動物祖先在細胞旌旗燈號轉導、基因表達調控模塊上特有的“立異”,使它的細胞獲得了跟著時空前提改變自身類型的能力,當作為干細胞的雛形。發育過程中,分歧細胞響應分歧的前提激活 / 封閉這些調控模塊,使得本身當作為某種特心猿意馬的細胞類型。
細胞堆積與分化呈現的時候不分先后,這兩個特點使動物的多細胞祖先得以闡揚群體內部多樣性的潛力,正如人群中個別的專性分工、普遍合作大大加速了人類社會的成長歷程,多細胞動物的進化速度也大大提高,最終迎來了 6 億年前的寒武紀生物大爆發。

* 把結論放在前面是為了賜顧幫襯那些其實火燒眉毛想知道謎底的伴侶,也許你們很忙沒有太多時候看完這么長的文字。但沒有關系,若是新的研究改變了這一結論,我會第一時候更新,以免你們記住的是錯誤的謎底。科學理論就是這樣,此刻看來信誓旦旦的工具指不心猿意馬哪天就變了,似乎讓人很沒有平安感。
* 不外仍是不由得想說,結論有些死板,但接下來藏在它背后的路程才真正出色。若是可以或許看到,一步一步嚴謹的科學研究中同化著時不時閃灼的靈光乍現,像聽著恢弘大氣又靈動活躍的交響樂,必然可以抹去那些干巴巴的理論帶來的不平安感,帶來心靈上的安慰,錯過了會很可惜的。
序:旅途起頭之前...
接下來可能是一段有些辛勞的路,在啟程之前,我們需要從題本家兒專心的問題描述中清算出幾個主要的行囊:
- 揣度進化汗青:進化生物學是一門汗青性學科,正如我們在這篇回覆中在做的這樣,它試圖說明生命的汗青以及造當作這些汗青事務的原因。可是發生的汗青已經遠去,我們看不到轉變是若何發生的,也不雅察不到是什么原因在起感化。化石凝固了時候,作為直接的證據可以告訴我們汗青中的那一刻是如何的;但更多時辰,我們必需經由過程不那么直接的證據來推演這些汗青以及原因。從進化的框架來看,萬萬種分歧的生物由獨自一個祖進步前輩化而來,那個祖先就是生命之樹的“根”。跟著時候長河的流動,一個物種可能會因一些性狀的分歧飾變而“分叉”——進化為兩個分歧物種。這樣的過程不竭反復,歷經時候大河的澆灌,孕育了現在多姿多彩的生物宿世界。那么,在我們今天可以或許不雅察到的生命之樹的結尾葉片即所有現存物種中,共享的性狀就應該是從第一個進化出這一特征的祖先那邊傳承下來的 (當然,趨同進化也可以塑造相似的性狀,于是區分來歷于配合祖先的相似性狀與自力趨同的相似性狀就當作為進化研究中必不成少的環節。具體方式有些復雜,這可所以別的一個故事了)。近緣物種分隔的時候短,共享的性狀就多一些;遠緣的物種分隔的時候長,保留的配合性狀少一些。若是某個特征只進化過一次并在所有的后裔中都保留了下來,那么這個共享的特征就可覺得這些分歧物種擁有配合祖先供給最好的證據。是以,我們可以經由過程比力分歧類群間的各類性狀特征來確定它們之間親緣關系的遠近,還可以借由那些祖源性狀描畫出它們配合祖先最可能的樣子。
- 用于進行進化揣度的性狀特征不僅僅限于外部形態學特征,還包羅行為、細胞布局、生化分子的布局以及染色體的布局。此刻最常用的證據是 DNA 序列,因為它在進化上相對不變、每個堿基位點的四個可能狀況不像其他表型特征輕易發生歧義,所以以它為按照構建的進化樹被認為是最靠得住的。
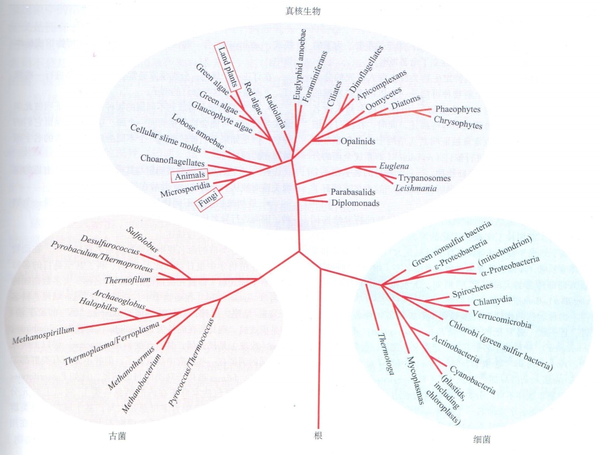
回到問題自己,
- “動物”一詞 (animals) 在現今的學術論文中專指動物總界 (Holozoa) 中的后活潑物 (Metazoa),詳參 @林十之 的回覆 地球上最初的生命為什么有的進化當作了植物有的進化當作了動物
- 原活潑物是在動物總界中單細胞個別的類群,固然在此中人們也不雅察到存在群居習性,但這樣的群體仍是由多個相對自力的細胞個別集聚而當作,每個細胞都具有營養、呼吸、分泌、生殖等作為一個動物所具有的一切機能。
- 動物總界中剩下的多細胞動物就是后活潑物,與原活潑物聚群底子的區別在于,細胞分化當作了分歧類型,各自執行專一的功能,互相依靠著構成動物個別而保存。在后活潑物中,一些類群因為特征上的不典型,與其他的動物不太相似,似乎有著更原始的特征,就被人們劃分出去,起名“側活潑物”(進化的側枝)、“中活潑物”(側枝與本家兒干之間的枝=_=),剩下的冠名“真后活潑物”。例如題本家兒提到的多孔動物,也就是那篇 Nature 文章的本家兒角海綿動物,仍是側活潑物獨一的現存門類,固然是多細胞動物,但身體只由兩層細胞構成,有怪異的水溝系統,在胚胎發育等方面也與其他多細胞生物顯著分歧。海綿動物的進化似乎一向處于相對障礙的狀況,現存的海綿動物與古老的海綿動物化石不同很小,它們應該是所謂的最“不在進化樹本家兒干”上的一類——直接被“側生”了。但不在本家兒干不代表它不主要,恰好因為海綿動物進化相對遲緩,保留了較多的古老特征,才給人們留下了絕好的追溯汗青的材料——用它和動物的原生親戚進行比力,我們甚至可以描畫出單細胞動物與多細胞動物分隔之初的那個配合祖先的肖像,進而推表演從單細胞祖先到多細胞動物的這段汗青中最可能發生了些什么。
- 再次保舉回覆 地球上最初的生命為什么有的進化當作了植物有的進化當作了動物 此中提到的基于 DNA 序列構建的系統發生闡發構建的進化樹展示了各類原活潑物與后活潑物的進化地位,而我們找到的兩個沖破口——海綿動物 (Porifera/sponge) 以及我們的單細胞親屬領鞭毛蟲 (Choanoflagellates),恰是諸多岔路中挨得比來的兩條。

1. 啟程:衣領、鞭毛、海綿
16 宿世紀末顯微鏡的發現在生物學的成長史中具有里程碑式的意義,人們終于可以做到“明察秋毫”,去不雅察生命更為底層的形式了。領鞭毛蟲初次被不雅察到是在三百年后,這種糊口在海水中自由游動的單細胞微生物剛一進入人們的視野就因為出格的表面形態令人們感覺倍感熟悉:一條很長的鞭毛,被一圈細胞膜概況的凸起物環抱,形當作一條圓柱狀的“衣領”。領鞭毛蟲是以而得名(Choanoflagellates, choano- 源于希臘語的“衣領”, flagellum 在拉丁語中意指鞭毛)。

這小細胞蟲好可愛啊,不外近似的細胞我身體里仿佛也有不少?(誤)
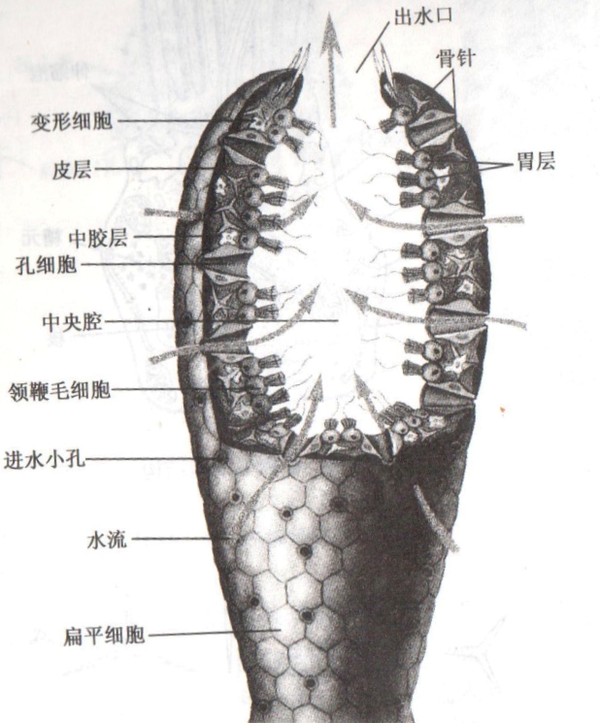
真正讓昔時的發現者感應熟悉的,是呈現在海綿動物體內的一種細胞類型——領鞭毛細胞 (choanocyte,簡稱領細胞)。
* 人們最早利用的海綿就是這些海綿動物,它們的皮層分布著很多孔洞,在水中糊口時供給水暢通過,海綿就用領細胞將經由過程的食物顆粒吞噬。因為這樣的多孔布局,海綿在打撈晾干后可以接收大量水分,可以被便利地利用。不外此刻日常所用的海綿都是用發泡材料建造的了。
領細胞布局與領鞭毛蟲極其相似,以至于發現者幾乎立即斷心猿意馬二者有著極近的親緣關系。有關更為詳盡的比力,詳參 @兔小灰 的回覆 海綿的領細胞和領鞭毛蟲在布局和功能上有什么區別
從細胞布局上看:
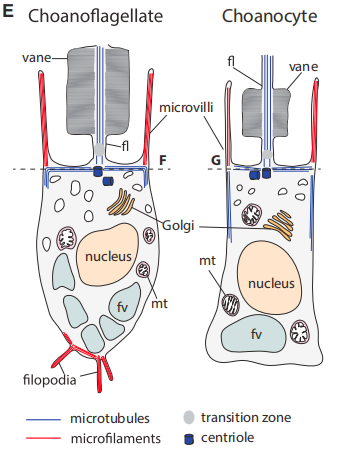
人們甚至認為,海綿動物中的領細胞就是由領鞭毛蟲進化而來的。之后研究者們又垂垂發現,近似這樣鞭毛 - 衣領的細胞形態,在動物界是半斤八兩遍及的:幾乎所有的動物,在個別發育某些階段或是部門細胞中,都可以或許不雅察到相似的布局 (想想那些小蝌蚪)。于是人們揣度,動物的祖先最初應該是領鞭毛蟲的那個樣子,然后聚團分化,一步一步往更復雜更高級的樣子成長。一百多年后,基于分子證據的系統發生研究終于確證了,領鞭毛蟲是原活潑物中離我們比來的門類,領鞭毛蟲與所有的后活潑物囊括了六億年前一個真核細胞 (被稱作 Urchoanozoan) 的所有后裔。同時,在后活潑物門類中,海綿動物是最原始、最古老的一類,比擬別的的動物,我們更有可能從海綿中發現配合祖先的影子。
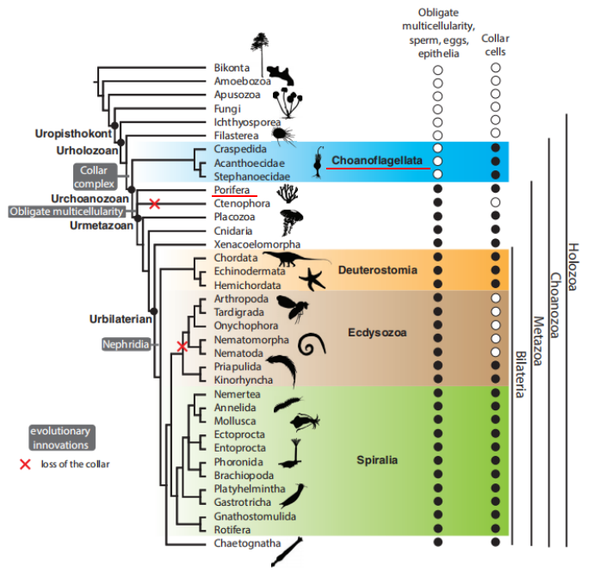
鞭毛 - 衣領布局的特征在領鞭毛蟲與后活潑物中被普遍共享,并且形當作這一布局的卵白質組分也大同小異,但在其他真核生物類群中,鞭毛的微布局就有著很大的差別。別的,人們在領鞭毛蟲的基因組上發現了很多編碼動物細胞微絨毛的同源基因,而且某些基因家族只在領鞭毛蟲與后活潑物中呈現,這些發現似乎都是對領鞭毛蟲與后活潑物來歷于配合祖先的有力證實。所以風行的不雅點是,動物的祖先就是有著領鞭毛特征的那個樣子,但在旅途的第 4 站就會看到,這個設法方才被證實是不合錯誤的。
接下來的問題是,原本自由安閑零丁游動的個別,又是若何攢聚到一路的呢?
2. 多細胞的發源:克隆相連
不需要任何專業常識,每小我都可以很輕易猜想到,從單個細胞釀成多個細胞無非兩種環境:細胞割裂的時辰沒有分手;或者多個分歧來歷的單細胞個別堆積黏連。放眼整個動物界,現存的所有動物都是經由克隆型的體例發育而來,即一個受精卵不竭割裂形當作多細胞布局,進而當作長為個別。從這一點來看,謎底似乎昭然若揭,動物的多細胞來歷應該就是第一種環境。可真的會是這樣嗎?
...
...
...
是的,就是這樣。有時辰幻化莫測的天然也會很誠懇,并沒有那么多轉折。不外,堆積型形當作策略也并未被其他生物遺棄,在天然界諸多門類中這種分歧于動物的多細胞連系體例經常能被不雅察到。
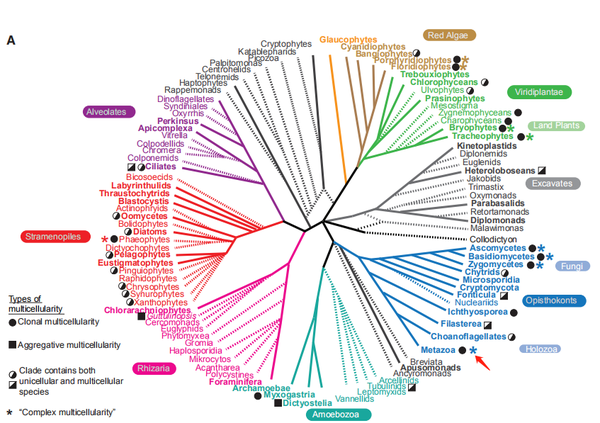
鑒于多細胞的糊口體例是如斯普遍地遍布整個天然界,且分歧門類的多細胞形當作體例在細節上存在諸多差別 (克隆型體例中還存在分歧的類型,但與我們故事本家兒線關系不大,有樂趣可以參閱參考文獻 [1] ), 學界傾標的目的于認為多細胞特征在進化過程中自力發生了很多次,不是多細胞生物的共有祖征。至于為什么動物祖先采用了這種而不是那種多細胞化的體例,真實的原因已無法確證,研究者們只能作一些可能的猜想:堆積型體例要求遺傳布景各不不異的細胞個別放下“自私”共同努力,且非論群體中的“變節者”可以或許半斤八兩輕易地粉碎群體布局,可是差別者的趨同也需要比克隆型更多的步調與前提。于是,進化出堆積型多細胞化體例的壁壘就比克隆型要高,前者也只能是特心猿意馬前提特心猿意馬選擇壓力下較為出格的產品了。
進一步的深切研究比力了大量多細胞化糊口的生物門類的基因表達,挖掘到了一系列與多細胞化相關的基因,所有的這些基因在功能上都可以歸為編碼胞外基質卵白與調控胞漿移動兩類。這暗示著,無論各類生物進化路徑有何等迥異,在多細胞化的分子機制上是趨同的:胞外基質黏連卵白與細胞割裂時的胞漿分派調控本家兒導了“細胞團”的形當作。那之后,二者繼續進化,一些通路上的漸變微調改變了細胞黏連的位置與割裂的偏向,付與原本圓滑的細胞團以極性,再加上時空特異的細胞類型分化,塑造了后活潑物多胚層的囊胚發育模式。
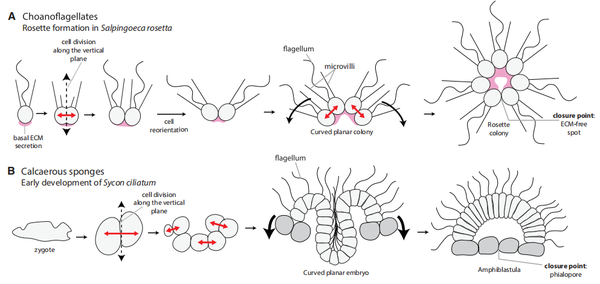
在將后活潑物、領鞭毛蟲的多細胞化體例與它們較近的類群 (上圖箭頭上方,Filasterea 蜷絲球蟲,Ichthyophonida 魚孢霉) 做比力時,后活潑物 / 領鞭毛蟲、蜷絲球蟲、魚孢霉輕易被區分為三類,每類之間有較大的的分歧。但動物與領鞭毛蟲之間是極其相似的,于是可以揣度,動物克隆型的多細胞化體例,在與領鞭毛蟲發生進化不合之前就已經呈現,動物的單細胞祖先該當是先營單細胞克隆形當作的群體糊口,再逐漸分化整合為多細胞個別的。
3. 細胞分化:流動著的樂團
形當作細胞團后,蜷絲球蟲、魚孢霉、領鞭毛蟲的孤傲心靈似乎已經獲得了知足,一向保留著個別在群體中的自力性,再也不肯多為鄰人支出些什么了。爾后活潑物是生成的革命者,它要操縱一切可以操縱的材料,一塊一塊搭建攀上復雜性岑嶺的階梯。
基于持久不雅察的證據,對于細胞分化的發源問題提出了兩種假說:
- 因為人們在不雅察各類單細胞生物的糊口史時已經發現,它們分歧糊口史期間對應著出格的細胞類型,響應地,這些細胞類型也對應著各自怪異的功能。例如,古蟲界的一些物種可以在變形蟲、鞭毛蟲、囊狀形態之間進行切換以順應分歧的保存需要;某些領鞭毛蟲如 Salpingoeca rosetta 也可以自由地改變自身的形態、功能以及糊口體例。是以,這種猜想認為,行使各類功能的諸多細胞類型對我們的祖先來說已經是現有的材料,即功能分化發生的時候早于多細胞化,動物祖先只是合理地放置這些負責分歧功能的細胞在合適的時候位置上呈現,一個未分化的細胞在某時達到了一個位置,它就將領受到調控分化的旌旗燈號,然后轉換當作那種需要的細胞類型。這被稱為 TST (Temporal-to-Spatial Transition, 時空轉換) 假說。
- 別的一方面,人們也看到,在原活潑物中由單個細胞完當作的進食、活動、分泌、生殖等功能,在后活潑物中被分歧的細胞所負責,似乎動物細胞顛末分化,功能越來越專精單一。于是,分歧的假設被提出了:細胞分化是在多細胞化之后發生,最初那個細胞集體中的每個當作員都在本身體內預載了周全的功能,跟著分化的進行,分歧的細胞丟失落了分歧的功能,朝著專一化成長,最終由這些功能專一的零部件組成個別。這是 DOL (Division of Labor, 功能不合) 假說。
固然對細胞分化的發源問題學界爭論已久,但這兩種假說并不是互斥的,動物祖先在進化過程中也有分階段采用兩種策略的可能。不外文章開首提到的今日頒發的 Nature 論文 (參考文獻 [2] ),為 TST 假說供給了很多證據,之后 TST 可能要占很長一段時候的優勢了。
4. 我們的祖先并不像小蝌蚪?
鑒于動物來自于領鞭毛蟲一樣的祖先這個由來已久的假設尚沒有被驗證、海綿領細胞與領鞭毛蟲同源這一猜想也因二者其實過分相像使人們一百多年來疏于尋找切當的證據,來自澳大利亞昆士蘭大學的 Bernand Degnan 傳授研究團隊具體比力了海綿三種分歧體細胞類型在基因表達上與領鞭毛蟲、蜷絲狀蟲、魚孢霉分歧糊口史階段的差別。
拔取領細胞之外的別的兩種海綿細胞是因為,人們在先期的研究中認為,那兩種細胞類型的呈現在進化汗青上早于領鞭毛蟲與動物配合祖先的分隔,別離對應到與魚孢霉、蜷絲狀蟲分隔的時候點。若是按人們之前認為的那樣,領細胞由古老的領鞭毛蟲進化而來,領細胞的細胞類型特征應該半斤八兩接近領鞭毛蟲。
細胞類型的劃分是經由過程區分細胞的“焦點調控復合體”的表達環境來實現 (參考文獻 [3] ),簡言之,近似于一排開關節制著一盞燈的顏色,最終燈光呈現出來的色彩由這些開關的組合狀況而決議。
成果很令人驚奇,領細胞在三種細胞類型中竟然是最“海綿”的那一個——它和其他的原活潑物有著最不相似的細胞類型,表達著最多海綿獨有的基因。也就是說,它這么像領鞭毛蟲并不是因為和領鞭毛蟲有著何等近的親緣關系,那個動物有著領鞭毛樣細胞團狀祖先的不雅點也就不當作立了。別的,此中一種海綿細胞在調控細胞增殖等“干細胞模塊”基因上有較高表達,而在其他原活潑物的滋生階段也不雅察到這些基因近似的表達傾標的目的,而這樣的表達模式與較為高檔的動物類群干細胞很是相似。
之后研究者用熒光染色追蹤發育過程中三種細胞,發現了它們在發育過程中存在遍及的互相轉化現象,細胞類型甚至可以往返改變。這供給了證據來拒絕 DOL 假說,動物祖先細胞的分化,應該更接近 TST 闡述的細胞類型隨時空前提改變而轉換的模式。
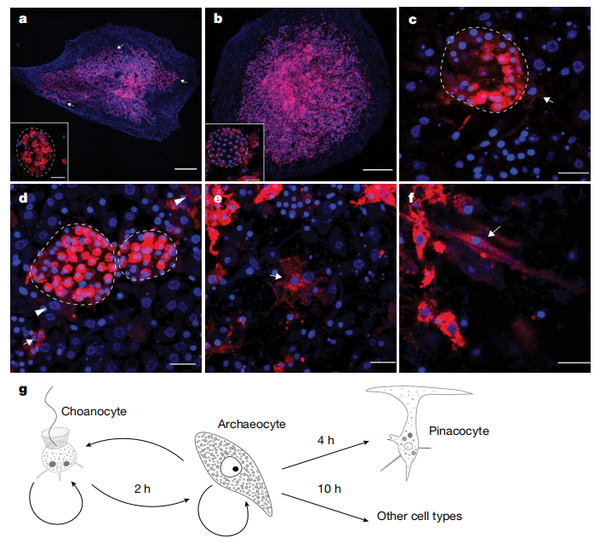
文章否認了那個我們來歷于領鞭毛蟲團團的不雅點,而且揣度動物祖先的多細胞狀況中每個細胞并不是一樣的,它們在半斤八兩早的時候段中已經呈現分化而且會跟著時空前提改變它們的細胞狀況。動物祖先基因組上旌旗燈號轉導通路、表達調控元件的改變,讓它學會了闡揚本身的多能性潛力,用緊密的調控系統實現了分歧狀況細胞的分化共存,進化當作了第一個多細胞動物。
從頭至尾聲:“丟掉的一環”
進化上的“為何”遠比“若何”難以回覆,因為汗青無法重演,我們難以確定生物類群的特征與其命運有著如何的因果聯系。動物中復雜度較低、現生門類較少的這些類群,并不是因為走上了進化的岔路、當作了生命史上的不測,它們之所所以它們,可能不需要有一個具體的原因。達爾文對生命之樹的描述富有詩意地蘊含了對待每一個物種平等的立場:
統一綱所有生物的親緣關系有時可以用一棵大樹來形容,我相信這一比方根基上是真切的。綠色的冒芽小細枝可以代表著現生物種,而那些前些年長出來的小枝則可代表連續串滅盡了的物種。在每一個發展期,所有當作長中的枝條都測驗考試標的目的各個偏向長出枝杈,并從其四周的細枝和枝杈中穿出甚至殺滅它們,恰如一個物種和一群物種在激烈的保存斗爭中所一向在進行的對其他物種的征服一樣。本家兒枝干分當作巨枝,巨枝再慢慢分當作越來越小的樹枝,而這些枝杈自己都曾是——當樹還小的時辰——冒芽的細枝;此刻的和舊日的芽間形當作的這種分杈關系可以或許很好地代表所有現生物種和已滅盡物種在分類中層層附屬的關系。在那些樹還尚矮小時長出的很多富強的細枝中,可能只有兩三條此刻長當作了大枝,而且不單幸存下來,還發生了其他的分枝;近似地,在曩昔漫長的地質期間所保存的物種中,只有少少數留下了依舊存活的變異了的后裔。從樹的最初發展起頭,很多本家兒枝和杈枝已經枯萎脫落了;這些脫落的巨細紛歧的樹枝可以代表那些整個目、科或者屬都沒有現生后裔的類群——他們僅以化石的形式為我們所知。就像我們偶然可以看到的那樣,一個細弱舒展的樹枝從一棵樹底部的樹杈上長出來,并因為某些偶爾的機遇受到偏心而且依然暢旺地存在世,于是我們有機遇看到像鴨嘴獸或南美肺魚這樣的動物;在些許水平上,它們毗連起生命的兩大分支,并且它們顯然因為糊口于一個呵護地而幸免于致命的競爭。因為芽經由過程發展再發生新芽,而若是這些新芽生命力興旺就會長當作枝條并蓋過四周很多較弱的枝條,是以我相信恢弘的生命之樹亦是代復一代地如斯:這棵生命之樹用它破落和枯死的樹枝填充了地殼,用它不竭分出的、斑斕的枝杈籠蓋了大地。
時候的長河雖一去不返,人類的雙眼卻可以借著思惟洞穿工夫,在腦中描畫出那個曩昔了的、看不到的,但我們相信著的真實宿世界。也許這恰是我們研究汗青、研究天然、研究宇宙的意義:我們可以憑著本身的能利巴握住涓滴的線索,還原我們不曾履歷的過往,指導我們正在度日的當下,燃起我們面臨未知的勇氣。
參考文獻
[1] Brunet, Thibaut, and Nicole King. "The origin of animal multicellularity and cell differentiation."Developmental cell 43.2 (2017): 124-140.
[2] Sogabe, Shunsuke, et al. "Pluripotency and the origin of animal multicellularity."Nature 570(2019): 519–522
[3] Arendt, Detlev, et al. "The origin and evolution of cell types."Nature Reviews Genetics 17.12 (2016): 744.
- 發表于 2019-07-09 00:22
- 閱讀 ( 1890 )
- 分類:其他類型
0 篇文章
作家榜 ?
-
 xiaonan123
189 文章
xiaonan123
189 文章
-
 湯依妹兒
97 文章
湯依妹兒
97 文章
-
 luogf229
46 文章
luogf229
46 文章
-
 jy02406749
45 文章
jy02406749
45 文章
-
 小凡
34 文章
小凡
34 文章
-
 Daisy萌
32 文章
Daisy萌
32 文章
-
 我的QQ3117863681
24 文章
我的QQ3117863681
24 文章
-
 華志健
23 文章
華志健
23 文章