幸好學習使我痛苦
 進修中感應疾苦該怎么辦?
進修中感應疾苦該怎么辦?
 vx: wenmiau
vx: wenmiau
熟悉這個問題,我們必需起首搞清晰,在認知 / 感知上,進修的素質是什么。
對于當作人來說,在一個自力進修的前后,我們熟悉宿世界的元問題體例和成果并沒有發生底子的改變,無論是圖像事物對象仍是聲學事物對象,我們感知的根本模式(邊、角、活動關系,頻率)沒有發生太多轉變。改變的是比力高級的聯系和認知成果,這本家兒要表現在記憶和邏輯兩個方面[1]. 無論是前者仍是后者,在更藐小的標準上,都表現為神經元之間毗連的形當作或重構(斷開和強化)。在宏不雅感知上,這個過程表現為“理解”。基于我們自身的體驗,進修表現為改變舊的熟悉模式和形當作新的認知模式,好比進修一種說話,舊有的字符組合變得有意義,形當作我們所領會的“單詞”。在大腦的剖解上,這對應為舊有的神經突觸毗連的斷開和新的突觸毗連的形當作以及強化。這些過程在大腦中可以以很快的速度發生,幾十分鐘的“進修”就能發生可不雅的大腦布局改變結果[2]:
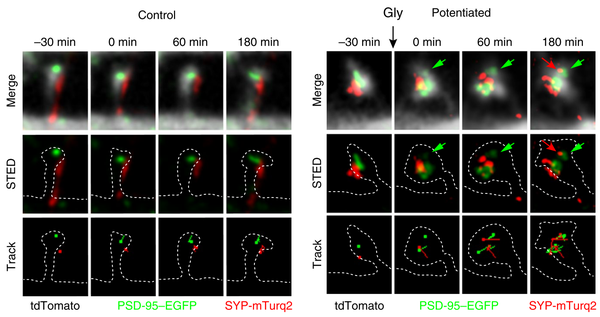
毗連的強化表示為兩個形式:1)更多神經元介入到統一個使命的激活突觸毗連中來[3],形當作協同放電;2)此中單個神經元的放電強度跟著進修過程獲得強化直至飽和。從這個意義上講,大腦是一種“數 - 模夾雜電路”。這樣的改變顛末數天 / 月 / 年,在剖解上甚至能表現到肉眼可見的標準上[4, 5],分歧專業技術者大腦布局是以會有顯著的宏不雅差別[6].
以上的過程中我們的感受是什么呢?對于這一點,我們當然可以類比于身體的熬煉:我們幾乎都知道熬煉身體對我們有益,可是在熬煉的過程中,大部門人會感受疾苦、委靡以及對意志的挑戰。不外這種類比不是從“第一性道理”[7]上作出的詮釋,從而缺乏足夠的說服力。
現實上,進修的疾苦感觸感染甚至可以從剛起頭進修就發生,是以這個感觸感染跟大腦的“委靡”的關系不是本家兒導性的。而是跟我們身體的一些“本能性”的工具相關,也就說,其原因根植于我們大腦的更深處(無論是邏輯意義上仍是現實剖解意義上)。
從上面臨進修素質的闡發可以看出,進修的過程需要“除舊迎新”。而“舊”的事物和認知讓我們感觸感染到舒適[8], 形當作所謂“comfort zone ”[9]. 而“新”的事物和認知則帶來不確定性和埋沒的危險的感觸感染,這是的我們的身體像面臨目生危險情況一樣釋放去甲腎上腺素(noradrenaline, NE)[10]。這直接導致全身血管的縮短以及嚴重、壓力和不安的感觸感染。
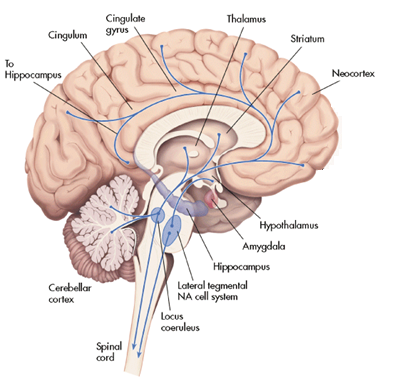
而對我們中的一些人來說,血管縮短自己就能導致四周的神經牽扯,從而導致神經性痛苦悲傷(詳見大腦沒有痛感神經那么頭痛是什么原因呢)[11].
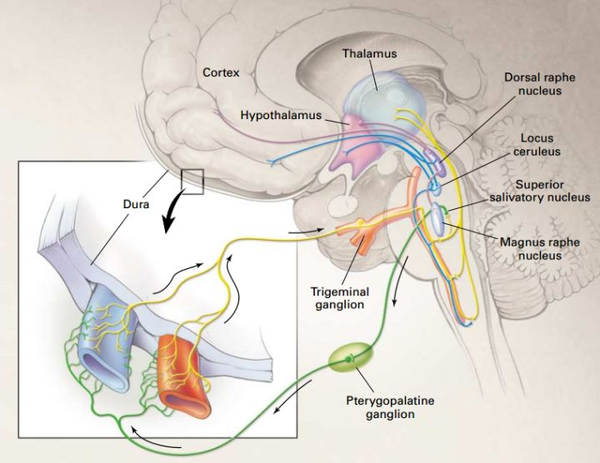
鬼使神差地,去甲腎上腺素 NE 也是一種神經遞質,在我們進修的過程中具有主要的感化,介入和調制進修的過程[12]. 是以它是我們進修過程的不雅測旌旗燈號之一[13]. 因為 NE 在進修過程中的雙重感化(介入進修過程神經調制 以及 發生壓力感觸感染),進修行為自己必然陪伴著壓力感等疾苦的感受。反之,若是進修過程中沒有這樣難熬難過的感受,則幾乎可以斷心猿意馬沒有真正的學到工具。當然,跟著進修的深切,“新”的工具逐漸被感知、熟悉和理解,我們的大腦會發生別致感。這陪伴這多巴胺的排泄[14, 15],是以多巴胺是進修過程中的另一個本家兒要旌旗燈號[13]. 當然,比擬于 NE,多巴胺是后效的——在進修的起頭階段,因為缺乏“別致”的熟悉,我們很少體味到其結果。
所以,若是進修的時候足夠長,以履歷疾苦 - 興奮這兩個階段的話,我們的身體現實上標的目的大腦同時傳遞了兩個互相矛盾的旌旗燈號——疾苦和舒適 / 興奮。 在這一矛盾的旌旗燈號輸入下,多巴胺排泄的結果會被嚴重減弱,我們的大腦感知到的興奮和獎賞從而也會被嚴重減弱,這會減弱我們的進修的價值的垂青性;陪伴著的是,我們大腦對疾苦的敏感性增添[16]. 此消彼長,其成果天然是疾苦占有了本家兒導性。
總結:
- 進修的素質是神經系統的重構,這個過程陪伴著神經內排泄的轉變
- 1 的轉變會導致血流環境轉變和神經牽扯,少數人是以會頭痛,也就是一進修就頭痛。這種頭痛具有必然的病理性
- 1 的轉變會導致疾苦的感觸感染,是以從心理性上講,進修過程自己必然是疾苦的
- 進修的結果會發生必然的回報性獎勵,讓我們感應歡快和興奮,但在進修過程的疾苦的陪伴下,這種獎勵的效能被大大降低,從而減弱我們的進修念頭
- 進修疾苦一般是正常心理現象
[1] Horn, G. (1985). Memory, imprinting, and the brain: An inquiry into mechanisms.
[2] Hruska, M., Henderson, N., Le Marchand, S. J., Jafri, H., & Dalva, M. B. (2018). Synaptic nanomodules underlie the organization and plasticity of spine synapses.Nature neuroscience,21(5), 671.
[3] Chen, H., Tang, A. H., & Blanpied, T. A. (2018). Subsynaptic spatial organization as a regulator of synaptic strength and plasticity.Current opinion in neurobiology,51, 147-153.
[4] Van Den Heuvel, M. P., Mandl, R. C., Kahn, R. S., & Hulshoff Pol, H. E. (2009). Functionally linked resting‐state networks reflect the underlying structural connectivity architecture of the human brain.Human brain mapping,30(10), 3127-3141.
[5] Parks, E. L., & Madden, D. J. (2013). Brain connectivity and visual attention.Brain Connectivity,3(4), 317-338.
[6] Gaser, C., & Schlaug, G. (2003). Brain structures differ between musicians and non-musicians.Journal of Neuroscience,23(27), 9240-9245.
[7] Irwin, T. (1989).Aristotle's first principles. Clarendon Press.
[8] Parker‐Rees, R. (2007). Liking to be liked: Imitation, familiarity and pedagogy in the first years of life.Early Years,27(1), 3-17.
[9] Shibata, T., Kim, J., Hoffman, D. M., & Banks, M. S. (2011). The zone of comfort: Predicting visual discomfort with stereo displays.Journal of vision,11(8), 11-11.
[10] Mason, S. T., & Iversen, S. D. (1975). Learning in the absence of forebrain noradrenaline.Nature,258(5534), 422.
[11] Tunis, M. M., & Wolff, H. G. (1954). Studies on headache: Cranial artery vasoconstriction and muscle contraction headache.AMA Archives of Neurology & Psychiatry,71(4), 425-434.
[12] Tully, K., & Bolshakov, V. Y. (2010). Emotional enhancement of memory: how norepinephrine enables synaptic plasticity.Molecular brain,3(1), 15.
[13] Harley, C. W. (2004). Norepinephrine and dopamine as learning signals.Neural plasticity,11(3-4), 191-204.
[14] Suri, R. E., Bargas, J., & Arbib, M. A. (2001). Modeling functions of striatal dopamine modulation in learning and planning.Neuroscience,103(1), 65-85.
[15] Wise, R. A. (2004). Dopamine, learning and motivation.Nature reviews neuroscience,5(6), 483.
[16] Talmi, D., Dayan, P., Kiebel, S. J., Frith, C. D., & Dolan, R. J. (2009). How humans integrate the prospects of pain and reward during choice.Journal of Neuroscience,29(46), 14617-14626.
- 發表于 2019-05-21 21:00
- 閱讀 ( 942 )
- 分類:其他類型
0 篇文章
作家榜 ?
-
 xiaonan123
189 文章
xiaonan123
189 文章
-
 湯依妹兒
97 文章
湯依妹兒
97 文章
-
 luogf229
46 文章
luogf229
46 文章
-
 jy02406749
45 文章
jy02406749
45 文章
-
 小凡
34 文章
小凡
34 文章
-
 Daisy萌
32 文章
Daisy萌
32 文章
-
 我的QQ3117863681
24 文章
我的QQ3117863681
24 文章
-
 華志健
23 文章
華志健
23 文章